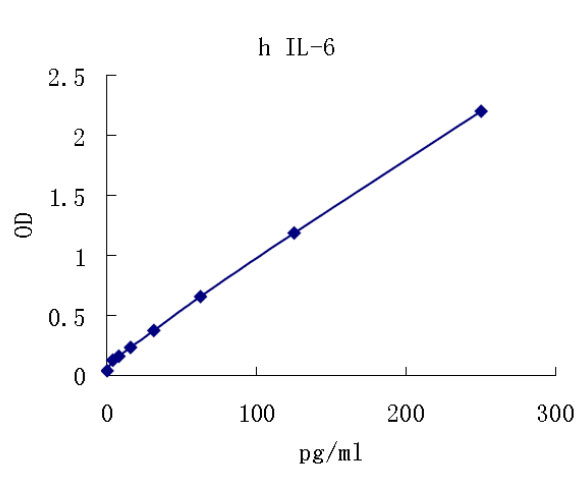
Interleukin 6(IL-6) is a multifunctional protein produced by lymphoid and non-lymphoid cells and by normal and transformed cells, including T cells, monocyte/macrophages, fibroblasts, hepatocytes, vascular endothelial cells, cardiac myxomas, bladder cell carcinomas, myelomas, astrogliomas and glioblastomas. The production of IL-6 in these cells is regulated, either positively or negatively, by a variety of signals including mitogens, antigenic stimulation, lipopolysaccharides, IL-1, TNF, PDGF and viruses. For reviews on IL-6, see references 1-5.
The human IL-6 cDNA sequence predicts a protein of 212 amino acid (aa) residues in length with two potential N-glycosylation sites. The hydrophobic N-terminal 28 aa residue signal peptide is cleaved to produce a mature protein of 184 amino acids with four cysteine residues and a predicted molecular mass of 21 kDa (6-9). The mouse IL-6 cDNA sequence shows a homology of 42% at the aa level when compared with the human sequence (10). On the basis of sequence similarity and gene structural motif similarity, IL-6 can be grouped in a family of cytokines that also includes OSM, G-CSF, LIF, and CNTF. All of these cytokines are predicted to have a four helix bundle structure similar to that found for growth hormone, suggesting that they all evolved from a common ancestral gene (11-13).
The effects of IL-6 on different cells are numerous and varied. The effect on B cells is stimulation of differentiation and antibody secretion (6, 14 - 17). IL-6 also affects T cells, acting as a co-stimulant with sub-optimal concentrations of PHA or Con A to stimulate IL-2 production and IL-2 receptor expression.
IL-6 exhibits growth factor activity for mature thymic or peripheral T-cells and reportedly enhances the differentiation of cytotoxic T-cells in the presence of IL-2 or IFN-γ (18-20). IL-6 stimulates production of acute phase proteins by hepatocytes (21) and has colony-stimulating activity on hematopoietic stem cells. IL-6 has growth factor activities and will stimulate the growth of myeloma/hybridoma/ plasmacytoma cells, EBV-transformed B cells, keratinocytes and mesangial cells (4, 5).
Kishimoto, T. et al. (1992) Science 258:5593.
Kishimoto, T. (1992) Int. Arch. Allergy Immunol. 99:172.
Hirano, T. et al. (1990) Immunol. Today 11:443.
Hirano, T. (1992) Clin. Immunol. Immunopathol. 62:S60.
Hirano, T. et al. (1990) in Peptide Growth Factors and their Receptors I, Sporn, M.B. and A.B.Roberts eds., Springer-Verlag, New York, p. 663.
Hirano, T. et al. (1986) Nature 324:73.
Haegeman, G. et al. (1986) Eur. J. Biochem. 159:625.
Zilberstein, A. et al. (1986) EMBO J. 5:2529.
May, L.T. et al. (1986) Proc. Natl. Acad. Sci. USA 83:8957.
Van Snick, J. et al. (1988) Eur. J. Immunol. 18:193.
Yasukawa, Y. et al. (1987) EMBO J. 6:2939.
Rose, T.M. and A.G. Bruce (1991) Proc. Natl. Acad. Sci. USA 88:8641d.
Bazan, J.F. (1991) Neuron 7:197.
Okada, M. et al. (1983) J. Exp. Med. 157:583.
Butler, L. et al. (1984) Proc. Natl. Acad. Sci. USA 81:2475.
Hirano, T. et al. (1984) J. Immunol. 133:798.
Kikutani, H. et al. (1985) J. Immunol. 134:990.
Lotz, M. et al. (1988) J. Exp. Med. 167:1253.
Tosato, G. and S.E. Pike (1988) J. Immunol. 141:1556.
Uttenhove, C. et al. (1988) J. Exp. Med. 167:1417.
Bauman, H. et al. (1984) J. Biol. Chem. 259:7331.