Intercellular Adhesion Molecule 1 (ICAM-1), also known as CD54, is a nearly ubiquitous transmembrane glycoprotein that plays a key role in leukocyte migration and activation (1, 2). Human ICAM-1 contains five Ig-like domains in its extracellular domain (ECD) and associates into non-covalently linked dimers (3, 4). Soluble forms of monomeric and dimeric ICAM-1 (sICAM-1) can be generated via proteolytic cleavage by cathepsin G, elastase, MMP-9, MMP-14/MT1-MMP, and TACE/ADAM17 (5 - 8). In the mouse, alternate splicing generates isoforms that lack particular Ig-like domains and are differentially sensitive to proteolysis (5). Within the ECD, human ICAM-1 shares 53% amino acid sequence identity with mouse and rat ICAM-1.
The principal binding partners of ICAM-1 are the leukocyte integrins LFA-1 (CD11a/CD18) and Mac-1 (CD11b/CD18) (9 - 11). The multivalency of dimeric ICAM-1 increases its strength of interaction with LFA-1 (9, 10). ICAM-1 also binds several non-integrin ligands including CD43/sialophorin, fibrinogen, hyaluronan, rhinoviruses, and Plasmodium falciparum-infected erythrocytes (12 - 16). At sites of inflammation, ICAM-1 is upregulated on endothelial and epithelial cells where it mediates the adhesion and paracellular migration of leukocytes expressing activated LFA-1 and Mac-1 (17 - 20). ICAM-1 ligation prolongs antigen presentation by dendritic cells and promotes T cell proliferation and cytokine release (21 - 23). ICAM-1 activation also participates in angiogenesis, wound healing, and bone metabolism (24 - 26).
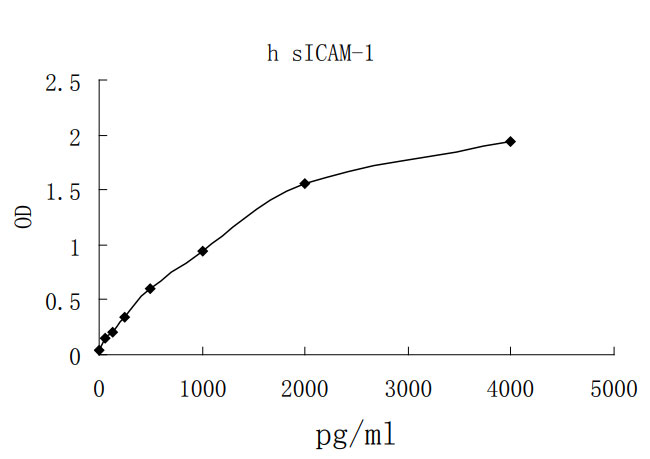
Soluble ICAM-1 has been reported in serum, cerebrospinal fluid, urine, and bronchoalveolar lavage fluid (2, 27 - 31). Elevated levels of sICAM-1 in these fluids are associated with cardiovascular disease, type 2 diabetes, organ transplant dysfunction, oxidant stress, abdominal fat mass, hypertension, liver disease, and certain malignancies (32 - 40). sICAM-1 promotes angiogenesis and serves as an indicator of vascular endothelial cell activation or damage (41, 42). It also functions as an inhibitor of transmembrane ICAM-1 mediated activities such as monocyte adhesion to activated endothelial cells and sensitivity of tumor cells to NK cell-mediated lysis (7, 8).
Hogg, N. et al. (1991) Chem. Immunol. 50:98.
Witkowska, A.M. and M.H. Borawska (2004) Eur. Cytokine Netw. 15:91.
Staunton, D.E. et al. (1988) Cell 52:925.
Reilly, P.L. et al. (1995) J. Immunol. 155:529.
Robledo, O. et al. (2003) Eur. J. Immunol. 33:1351.
Sithu, S.D. et al. (2007) J. Biol. Chem. 282:25010.
Fiore, E. et al. (2002) Oncogene 21:5213.
Tsakadze, N.L. et al. (2006) J. Biol. Chem. 281:3157.
Jun, C.D. et al. (2001) Proc. Natl. Acad. Sci. 98:6830.
Miller, J. et al. (1995) J. Exp. Med. 182:1231.
Diamond, M.S. et al. (1990) J. Cell Biol. 111:3129.
Rosenstein, Y. et al. (1991) Nature 354:233.
Pluskota, E. and S.E. D